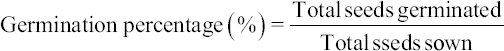1. Akinyele AO. Silvicultural Requirements of Seedlings of Buchholzia coriacea Engl. Ph. D Thesis Submitted to the Forest Resources Management, University of Ibadan;2007. 179.
2. Rom CR. Light threshold for apple tree canopy growth and development. Hortic Sci 1991;26:889-992.
3. Fenner M, Thompson K. The Ecology of Seeds. Cambridge, UK:Cambridge University Press;2005.
4. Simão E, Takaki M. Effect of light and temperature on seed germination in Tibouchina mutabilis (Vell.) Cogn. (Melastomataceae). Biota Neotrop 2008;8:63-8.
5. Guo C, Shen Y, Shi F. Effect of temperature, light, and storage time on the seed germination of Pinus bungeana Zucc. ex Endl:The role of seed-covering layers and abscisic acid changes. Forests 2020;11:300.
6. Hart JW. Light and Plant Growth. London:Unwin Ltd.;1988. 213.
7. Baskin CC, Baskin JM. Seeds:Ecology, Biogeography, and Evolution of Dormancy and Germination. 2nd ed. San Diego, CA:Academic Press;2014.
8. Fenner M. Germination tests of thirty-two East African weed species. Weed Res l980;20:135-8.
9. Muhanguzi HD, Obua J, Oryeni-Origa H. Influence of light quality on the germination characteristics of seeds of selected pioneer, understorey and canopy tree species in Kalinzu forest reserve, Uganda. Uganda J Agric Sci 2002;7:25-30.
10. Rojas-Aréchiga M, Mandujano MC, Golubov JK. Seed size and photoblastism in species belonging to tribe Cacteae (Cactaceae). J Plant Res 2013;126:373-86.
11. Ribeiro LC, Borghetti F. Comparative effects of desiccation, heat shock and high temperatures on seed germination of savanna and forest tree species. Austral Ecol 2014;39:267-78.
12. Souza ML, Fagundes M. Seed predation of Copaifera langsdorffii (Fabaceae):A tropical tree with supra-annual fruiting. Plant Species Biol 2017;32:66-73.
13. Veloso AC, Silva PS, Siqueira WK, Duarte KL, Gomes IL, Santos HT, Fagundes M. Intraspecific variation in seed size and light intensity affect seed germination and initial seedling growth of a tropical shrub. Acta Bot Brasilica 2017;31:736-41.
14. Seo M, Hanada A, Kuwahara A, Endo A, Okamoto M, Yamauchi Y, et al. Regulation of hormone metabolism in Arabidopsis seeds:Phytochrome regulation of abscisic acid metabolism and abscisic acid regulation of gibberellin metabolism. Plant J 2006;48:354-66.
15. Nonogaki H. Seed biology updates-highlights and new discoveries in seed dormancy and germination research. Front Plant Sci 2017;8:524.
16. Kołodziejek J, Patykowski J, Wala M. Effect of light, gibberellic acid and nitrogen source on germination of eight taxa from dissapearing European temperate forest, Potentillo albae-Quercetum. Sci Rep 2017;7:13924.
17. Kyereh B, Thompson J. Effect of light in the germination of forest trees in Ghana J Ecol 1999;87:772-83.
18. Gutterman Y. Strategies of seed dispersal and germination in plants inhabiting deserts. Bot Rev 1994;60:71-80.
19. ISTA. International Rules for Seed Testing. Zurich:The International Seed Testing Association;1996. 17.
20. Schelin M, Tigabu M, Eriksson I, Sawadogo L. Effect of scarification, gibberellic acid and dry heat treatments on the germination of Balanites aegyptiaca seeds from the Sudanian savanna in Burkina Faso. Seed Sci Technol 2003;31:605-17.
21. Bowers J. Does Ferocactus wislizeni (Cactaceae) have a between year seed bank?J Arid Environ 2000;45:197-205.
22. Oyedeji OF, Amadi JO, Sowunmi IL, Adebusuyi GA, Dunmade YA, Agboola RO. Influence of light on seed germination of Dialium guineense Willd. J Sustain Environ Manag 2018;10:63-72.
23. Flores J, González-Salvatierra C, Jurado E. Effect of light on seed germination and seedling shape of succulent species from Mexico. J Plant Ecol 2016;9:174-9.
24. Islam MO, Matsu S, Ichihashi S. Effects of light quality on seed germination and seedling growth of Cattleya orchids in vitro. J Jpn Soc Hortic Sci 1999;68:1132-8.
25. Rojas-Aréchiga M, Casa A, Vázquez-Yanes C. Seed germination of wild and cultivated Stenocereus stellatus (Cactaceae) from the Tehuacán-Cuicatlán Valley, Central México. J Arid Environ 2001;49:279-87.
26. Pearson TR, Burslem DF, Mullins CE, Dalling JW. Functional significance of photoblastic germination in neotropical pioneer trees:A seed's eye view. Funct Ecol 2003;17:394-402.
27. Ranieri BD, Lana TC, Negreiros D, Araújo LM, Fernades GW. Seed germination of Lavoisiera cordata Cogn. and Lavoisiera francavillana Cogn. (Melastomataceae), sympatric plant species from Serra do Cipó, Brazil. Acta Bot Bras 2003;17:523-30.
28. Godoi S, Takaki M. Effects of light and temperature on seed germination in Cecropia hololeuca Miq. (Cecropiaceae). Braz Arch Biol Technol 2004;47:185-91.
29. Benitez-Rodriguez JL, Orozco-Segovia A, Aréchiga MR. Light effect on seed germination of four Mammillaria species from the Tehuacán-Cuicatlán Valley, central México. Southwest Nat 2004;49:11-7.
30. Simão E, Socolowski F, Takaki M. The epiphytic Cactaceae Hylocereus setaceus (Salm-Dick ex DC.) Ralf Bauer seed germination is controlled by light and temperature. Braz Arch Biol Technol 2007;50:655-62.
31. Costa A, Dias AS, Grenho MG, Dias LS. Effects of dark or of red, blue or white light on germination of subterranean clover seeds. Emirates J Food Agric 2016;28:853-64.
32. Rakhmawati SU, Rahmadiyanto AN. The Effects of Light Color on Seed Germination of Markhamia stipulata (Wall.) Seem in ICBS Conference Proceedings, International Conference on Biological Science 2015, KnE Life Sciences;2017. 233-40.
33. Serrano-Bernardo F, Rosúa JL, Díaz-Miguel M. Light and temperature effects on seed germination of four native species of Mediterranean high mountains (Spain). Int J Exp Bot 2007;76:27-38.
34. Maloof JN, Borevitz JO, Weigel D, Chory J. Natural variation in phytochrome signaling. Semin Cell Dev Biol 2000;11:523-30.
35. Baskin CC, Baskin JM. Germination ecophysiology of herbaceous plant species in a temperature region. Am J Bot 1988;75:286-305.
36. Onyekwelu JC, Stimm B, Mosandl R, Olusola JA. Effects of light intensities on seed germination and early growth of Chrysophyllum albidum and Irvingia gabonensis seedlings. Niger J Forestry 2012;42:58-67.