![]()
Changes across time post-feed offer in the composition of total mixed ration for finishing beef cattle
Md. Ashraful Islam*
Department of Dairy Science, Faculty of Animal Science and Veterinary Medicine, Patuakhali Science and Technology University, Barishal, Bangladesh
ABSTRACT
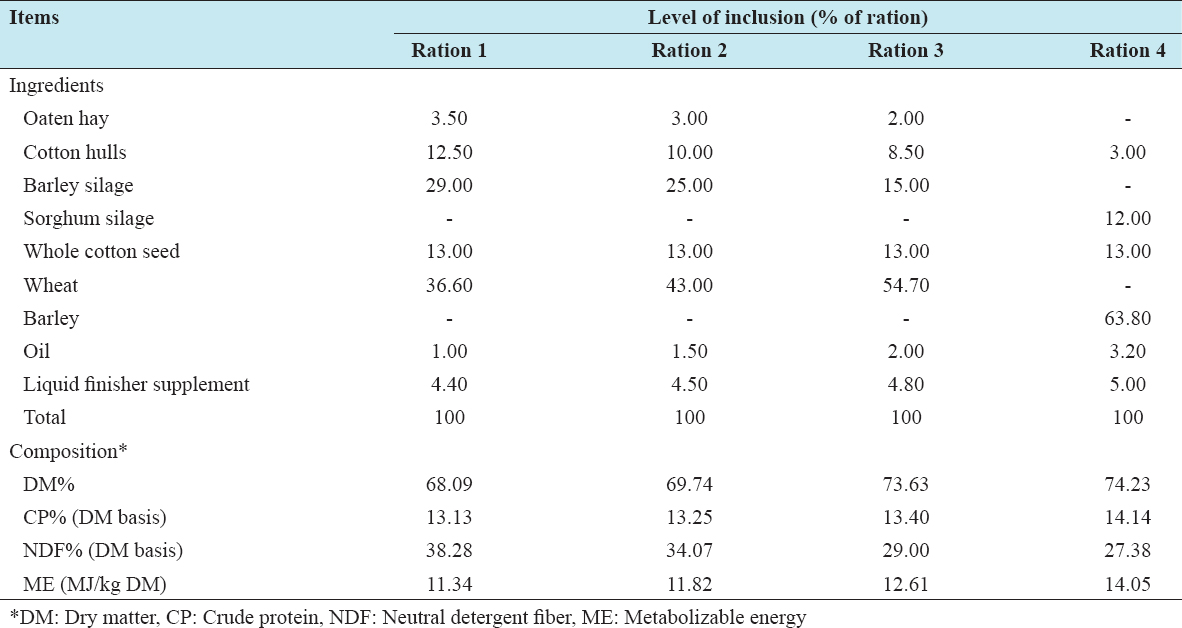
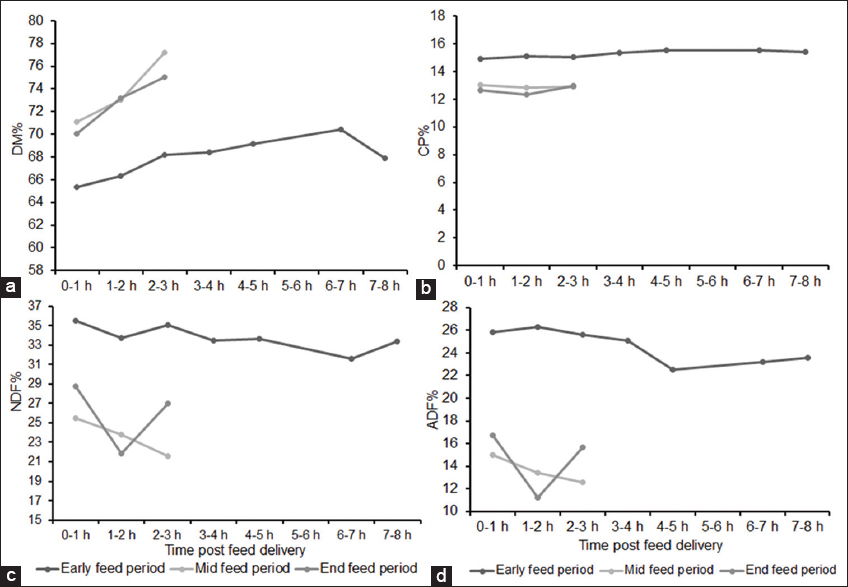
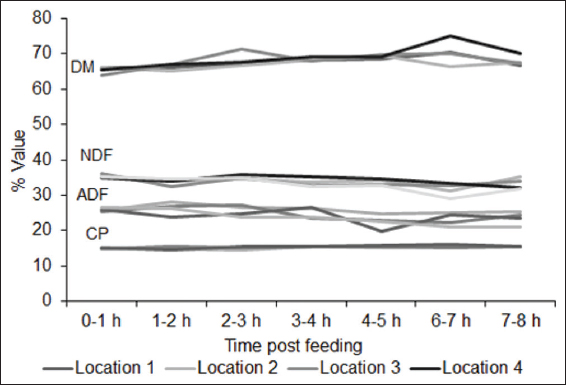
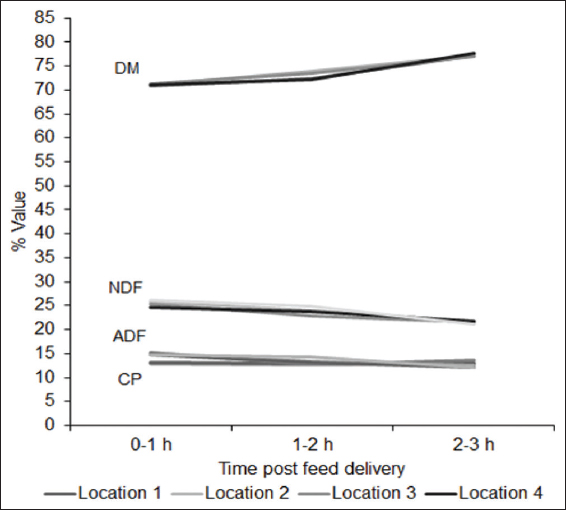
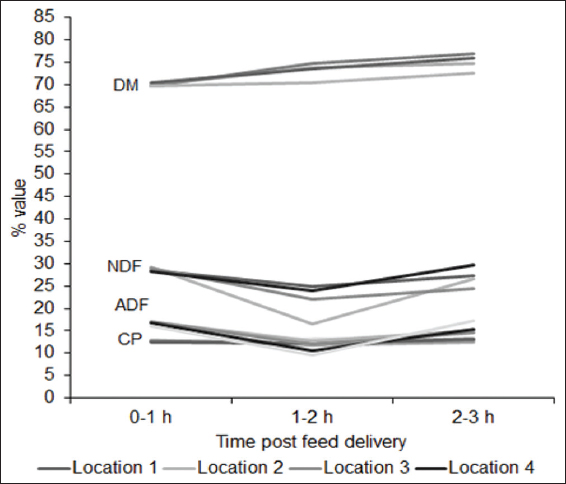
Cattle can select feed ingredients based on the physiological need and composition of ration provided. However, there is a paucity of knowledge regarding impact of selective feeding behavior of finishing beef cattle on the overall composition of the provided ration across hour of day during full feed period in the feedlot. Our objectives were to determine the impact of time post-feeding, location within the feed bunk, and their interaction on composition of the remaining feed. For this experiment, feed samples were collected during early- (days 3–5), mid- (days 41 and 42), and end- (days 71 and 72) feed periods from feed bunk of a commercial feedlot pen of 100 cattle gradually adapted to a high grain total mixed ration through transitions of four different rations. Samples were collected immediately after feed delivery and at different intervals of post-feed delivery from four equally spaced locations within the feed bunk and analyzed for dry matter (DM)%, crude protein (CP)%, acid detergent fiber (ADF)%, and neutral detergent fiber (NDF)% with an aim to look into changes in the composition of remaining feed in the feedlot bunk across time and location. For all three sampling periods, DM% and NDF% of feed differed significantly (P < 0.001) with an overall tendency to increase in DM% and a decrease in NDF% during early hours post-feeding, indicating selection of succulent and high-fiber feed. Early feed CP% differed significantly (P = 0.023) overtime but differences were non-significant for mid-feed (P = 0.400) and end-feed (P = 0.059) periods. Changes in ADF% were non-significant (P = 0.074) for early-feed period but were significant (P < 0.001) for mid- and end-feed periods. Locations within the feed bunk had no significant effect on composition of feed. Interactions between time post-feeding and locations were found significant (P = 0.011) only for end-feed NDF%. Overall these results suggest feedlot cattle select feed primarily for fiber and that affect composition of feed overtime. Given the differences in individual eating time and duration, multiple delivery of feed across 24 h and increasing feeding space can minimize the impact of selective feeding related altered composition of feed offered.
Keywords: Feed composition, feed sorting, feedlot cattle, neutral detergent fiber